
 Im Muskelfleisch von Rehen variierte die Cäsium-137 Aktivität jedes Jahr ausgesprochen saisonal, mit niedrigen Werten im Frühjahr und deutlich höheren Werten im Herbst. In der Abbildung 1 ist der Verlauf der Cs-137 Aktivität in Rehwild aus Bodenmais, von 1987 bis 2001, anhand der Monatsmediane gezeigt. Die Unterschiede zwischen der minimalen Kontamination im Frühjahr und der maximalen im Herbst sind erheblich und betragen eine Größenordnung. Im Gegensatz zu Rehen ist die Cäsium-137 Kontamination bei Wildschweinen deutlich weniger saisonal ausgeprägt.
Im Muskelfleisch von Rehen variierte die Cäsium-137 Aktivität jedes Jahr ausgesprochen saisonal, mit niedrigen Werten im Frühjahr und deutlich höheren Werten im Herbst. In der Abbildung 1 ist der Verlauf der Cs-137 Aktivität in Rehwild aus Bodenmais, von 1987 bis 2001, anhand der Monatsmediane gezeigt. Die Unterschiede zwischen der minimalen Kontamination im Frühjahr und der maximalen im Herbst sind erheblich und betragen eine Größenordnung. Im Gegensatz zu Rehen ist die Cäsium-137 Kontamination bei Wildschweinen deutlich weniger saisonal ausgeprägt.
Ursachen der Kontamination
Je nach Vegetationszustand und Zeitpunkt des Wintereinbruchs, fressen sich Rehe, von Ende September bis November ihren “Winterspeck” an, nehmen also wesentlich mehr Nahrung auf als sonst, hauptsächlich Blätter von Farnen, Himbeere, Brombeere, Heidelbeere und Pilze. Dornfarn und einige Pilzarten sind gerade dann relativ hoch mit Cs-137 belastet, also erreicht auch die Kontamination von Rehwild, durchschnittlich im Oktober/November ihren höchsten Stand.
Mit dem ersten Schneefall geht der Nahrungsbedarf der Rehe schlagartig zurück und sinkt, aufgrund des nun verminderten Stoffwechsels während des Winters bis fast auf die Hälfte der Menge ab, die im Oktober aufgenommen wird.
Da die biologische Halbwertzeit von Cäsium-137 in Rehen, mit ca. 10 Tagen relativ klein ist, nimmt auch die Kontamination der Tiere, mit Beginn des Schneefalls, schnell ab. Im Winter liegt in Bodenmais i.d.R. mehr als 1 Meter Schnee. Rehe nehmen dann den Großteil ihrer Nahrung an Winterfütterungen auf, wo nur gering kontaminiertes Futter angeboten wird.
Saisonales Muster der radioaktiven Kontamination bei Rehen
Zusätzlich fressen sie noch geringe Mengen Knospen von Sträuchern und jungen Bäumen, die aus dem Schnee herausragen, bzw. andere Waldpflanzen, die sie frei scharren. Knospen sind i.d.R. geringer belastet als die Blätter der meisten Waldpflanzen während der Vegetationsperiode. In Folge dessen bleiben die Cäsium 137 Gehalte im Muskelfleisch der Rehe den ganzen Winter über niedrig. Im Frühjahr beginnt mit der Aufnahme der ersten, relativ Cs-137 haltigen Waldbodenpflanzen, im Cs-Stoffwechsel von Rehwild ein, dem Winter entgegengesetzter Zyklus: Der Cs-137 Input durch die Nahrung nimmt zu, wodurch das Nuklid in der Muskulatur angereichert wird (Abbildung 1). Im Vergleich zum Rehwild ist die radioaktive Cäsium Belastung bei Rothirschen deutlich weniger saisonal.

Abb. 1: Cs-137 Aktivität in Rehen aus Bodenmais, Mai 2000 bis Mai 2001 (n=73)
Dieses saisonale Muster der Kontamination zeigte sich bei unseren Untersuchungen in jedem Jahr. In der Abbildung 1 ist der Verlauf der Cs-137 Aktivität in Rehwild aus Bodenmais, von 1987 bis 2001, anhand der Monatsmediane gezeigt. Die Unterschiede zwischen der minimalen Kontamination im Frühjahr und der maximalen im Herbst sind erheblich und betragen eine Größenordnung.
Von 1987 bis 2001 nahm die Cs-137 Aktivität in Rehen ab. Der höchste Medianwert wurde im Oktober 1987, mit 7.040 Bq/kg festgestellt. 1993 betrug der höchste Monats-median im Oktober 3.810 Bq/kg, 1999 im September noch 1.220 Bq/kg. 2000 erfolgte ein deutlicher Anstieg der Kontamination, mit 3.370 Bq/kg wurde im November 2000 der höchste monatliche Median seit 1993 festgestellt.
Zeitliche Entwicklung der Cäsium 137 Kontamination
In der Abbildung 2 ist die zeitliche Entwicklung der radioaktiven Belastung von Rehen anhand der Monatsmediane von Cäsium 137 für das Untersuchungsgebiet Bodenmais dargestellt.

Abb. 2: Zeitverlauf von Cs-137 in Rehfleisch aus Bodenmais. Monatsmediane von 1987 – 2003 (n=1.557)
Von 1987 bis 2001 nahm die Cs-137 Aktivität in Rehen ab. Der höchste Medianwert wurde im Oktober 1987, mit 7.040 Bq/kg festgestellt. 1993 betrug der höchste Monatsmedian im Oktober 3.810 Bq/kg, 1999 im September noch 1.220 Bq/kg.
2000 erfolgte ein deutlicher Anstieg der Kontamination, mit 3.370 Bq/kg wurde im November 2000 der höchste monatliche Median seit 1993 festgestellt.
Stoffwechsel und Kintetik
Der Radiocäsiumgehalt, der sich im Muskelfleisch der Rehe einstellt, wird, neben der Kinetik des Nuklids im Tierkörper wesentlichen von der mit der Nahrung aufgenommenen Aktivität des Nuklids bestimmt. Dieser Cs-137 Input wird durch die Größen
- 1. Nahrungsmenge pro Zeiteinheit und
- 2. Aktivitätsgehalt der Nahrungspflanzen
gesteuert. Beide Variablen werden wiederum durch andere Faktoren wie Klima, Standortverhältnisse etc. beeinflußt. Im folgenden sollen die Einflüsse dieser Variablen diskutiert werden:
Nahrungsmenge
Rehe nehmen im Jahreslauf unterschiedlich viel Nahrung zu sich, im Winter z.B. grundsätzlich weniger als im Sommer. Dieses Verhalten ist als stoffwechselphysiologische Anpassung an das im Winter verringerte Nahrungsangebot anzusehen.
Im Rahmen der vorliegenden Arbeit war es nicht möglich die Nahrungsmenge, die von Rehwild in den Untersuchungsgebieten aufgenommen wird, zu ermitteln. Daher wird in diesem Punkt auf den Fütterungsversuch von ELLENBERG (1978) zurückgegriffen, dessen Ergebnisse i.w. übertragbar sind.
Nahrungsverhalten im Gehege
Der Autor untersuchte das Nahrungsverhalten von 10 Rehböcken in einem Gehege, unter naturnahen Bedingungen. Er stellte fest, daß die Tiere, trotz konstanter Haltungsbedingungen und Zusammensetzung des Futters, im Jahresablauf unterschiedlich viel Nahrung aufnahmen (Abbildung 3). Besonders niedrig war die durchschnittliche Nahrungsaufnahme während der Brunft Ende Juli/Anfang August und zur Zeit des intensiven Geweihschubs im Januar und Anfang Februar. Schneefall, wenn auch nur in Spuren, führte ebenfalls zu einer verminderten Nahrungsaufnahme. Im Durchschnitt wurde die größte Futtermenge im Oktober aufgenommen. Da juvenile Rehe noch in der Wachstumsphase sind und i.d.R. nicht am Brunftgeschehen teilnehmen, wird sich ihre Nahrungsaufnahme von der adulter Tiere unterscheiden.
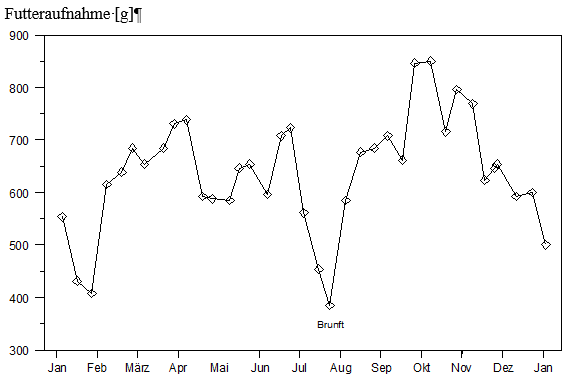
Abb. 3: Durchschnittliche Futteraufnahme (Gramm Kraftfutter pro Tier und Tag bei 88 % Trockensubstanz) von 10 drei- bis vierjährigen Rehböcken, verändert nach ELLENBERG (1978)
Nach den Angaben von DROZDZ und OSIECKI (1973), die einen Fütterungsversuch an
3 Rehen durchführten, nahmen Rehe im Sommer durchschnittlich 55O g, im Winter dagegen nur 350 g Nahrung (TS) pro Tag auf.
Nahrungsspektrum der Rehe aus Bodenmais
Rehe ernähren sich i.w. von leicht verdaulichen, nährstoffreichen Pflanzen, bevorzugt werden Blätter von Kräutern, Sträuchern und jungen Bäumen (BUBENIK 1984, ELLENBERG 1978, KLÖTZLI 1965).
Da die Zusammensetzung der Vegetation regional verschieden ist, variiert auch das Nahrungsspektrum der Rehe. Deshalb wurde das Nahrungsspektrum von Rehwild aus dem Untersuchungsgebiet Bodenmais genau untersucht. Auf die Ergebnisse soll im folgenden näher eingegangen werden.
Magenanalysen: Analyse von 122 Pansen
Untersucht wurden die Panseninhalte von 122 Rehen, nach der Methode von CHAMRAD und BOX (1964), die 1987 – 1990, jeweils von Juni bis Dezember in Bodenmais erlegt wurden. Die Arbeiten wurden von ALBERS (1992) durchgeführt. Die Methode beruht darauf, daß die im Pansen vorhandenen Pflanzenfragmente anhand einer Merkmals-kombination ihrer Epidermisstrukturen, jeweils bis zur Art bestimmt werden können (die Blattgewebe werden zwar durch die Magensäfte vorverdaut und in ihrer Struktur verändert, nicht aber deren kutinhaltige obere Abschlußhäute, die sich durch die Größe und Anordnung verschiedener Zellarten, Härchen sowie Wachsschichten voneinander unterscheiden und dadurch i.d.R. typisch für jede Pflanzenart sind).
Methode
Aus jedem Pansen wurden nach einem statistischen Verfahren 3 x 100 Pflanzenfragmente ausgewählt und möglichst bis zur Art bestimmt. Aus diesen Daten wurde der Prozentanteil, mit dem eine Pflanzenart im Mageninhalt vorkommt, errechnet (ALBERS 1990). Insgesamt wurden 114.000 einzelne Pflanzenfragmente unter dem Mikroskop bestimmt. Die Ergebnisse sind in Tabelle 1 dargestellt.
Tab. 1: Durchschnittliches Vorkommen (X) einer Pflanzenart im Pansen von Rehen
(n = 114) aus Bodenmais von Juni – Dezember, 1987 – 1990. F = Anteil an der Gesamtzahl der Pansen in denen eine Pflanzenart nachgewiesen wurde
| Pflanzenart | X [%] | F [%] |
| Himbeere | 17 | 77 |
| Farn | 13 | 76 |
| Brombeere | 12 | 34 |
| Eberesche | 8 | 70 |
| Heidelbeere | 8 | 64 |
| Hasenlattich | 6,2 | 34 |
| Fütterung | 5 | 18 |
| Pilze | 3,3 | 37 |
| Früchte | 2 | 33 |
| Rinde / Knospen | 1 | 39 |
| Gras | 0,7 | 24 |
| Pflanzenarten mit Anteil < 1 % | 17 | 95 |
| Pfl.fragmente nicht bestimmbar | 6,6 | 90 |
Eine Auflistung aller ermittelten Nahrungspflanzen ist in Tabelle IX im Anhang gegeben. Pflanzenfragmente von Farnen konnten unter dem Mikroskop nicht bis zur Art bestimmt werden und sind deshalb unter dem Begriff „Farne“ angegeben. Aufgrund der Beobachtungen im Untersuchungsgebiet an verbissenen Farnpflanzen, ist davon auszugehen, daß Dornfarn, Wald-Frauenfarn und Bergfarn von Rehen gefressen werden, während Adlerfarn nicht angerührt wird.
Allein die Angabe über das durchschnittliche Vorkommen einer Pflanzenart in allen untersuchten Pansen läßt noch keinen Schluß über deren Bedeutung für die Rehpopulation zu: wenn z.B. einzelne Tiere ausschließlich eine Pflanzenart gefressen haben, kann das einen geringen bis mäßigen Genuß in der Grundgesamtheit vortäuschen. Daher ist es wichtig, auch die Häufigkeit des Vorkommens (Frequenz) in den Pansen zu berücksichtigen. So deutet bei der Brombeere der relativ hohe X-Wert von 12 %, bei einer Frequenz von 34 % darauf hin, daß diese Pflanzenart in nur einem Drittel aller Pansen vorkam, allerdings zu einem relativ hohen Anteil. Ziemlich gleich war dagegen in allen Pansen der Anteil der Pflanzfragmente, der nicht bestimmbar war: in 90 % aller Pansen waren dies durchschnittlich 6,6 %.
39 Pflanzenarten wurden nachgewiesen
Insgesamt wurden 39 Pflanzenarten in der Nahrung nachgewiesen, wovon aber nur wenige Arten mengenmäßig von Bedeutung waren: durchschnittlich war der Anteil am Nahrungs-spektrum bei Himbeere mit 17 % am höchsten, gefolgt von Farn mit 13 % und Brombeere mit 12 %. Im mittleren Wertebereich lagen die Anteile von Eberesche und Heidelbeere mit je 8 %, sowie Hasenlattich mit 6,2 %.
Pflanzenbestandteile wie Heu, Äpfel etc. die aus der Winterfütterung stammten, machten durchschnittlich 5 % der Panseninhalte aus. Dieser Anteil ist, auf das ganze Jahr bezogen, sicherlich höher, denn aus den Monaten Januar bis Mai, wo die Rehe i.d.R. relativ viel Nahrung an Fütterungen aufnehmen, konnten keine Pansen untersucht werden.
Eiweißreiche und leicht verdauliche Pflanzenkostwird bevorzugt
Beachtenswert ist, daß in 95 % aller untersuchten Pansen viele verschiedene Pflanzenarten bestimmt wurden, deren jeweiliger Anteil kleiner als 1 % war. Nicht überraschend war degegen, daß Gras nur den unbedeutenden Anteil von 0,7 % am Nahrungsspektrum hatte. Diese Ergebnisse stimmen mit denen anderer Untersuchungen überein (o.g. Literatur), nach denen Rehe eiweißreiche und leicht verdauliche Pflanzenkost bevorzugen. Rothirsche nehmen im Gegensatz dazu z.B. relativ mehr zellulosehaltige Pflanzennahrung auf, wozu auch Gras zählt.
Die Zusammensetzung der Nahrungsbestandteile in den untersuchten Rehpansen variierte im Jahresverlauf beträchtlich. Die Zusammenhänge sind in der Abbildung 4 wiedergegeben.
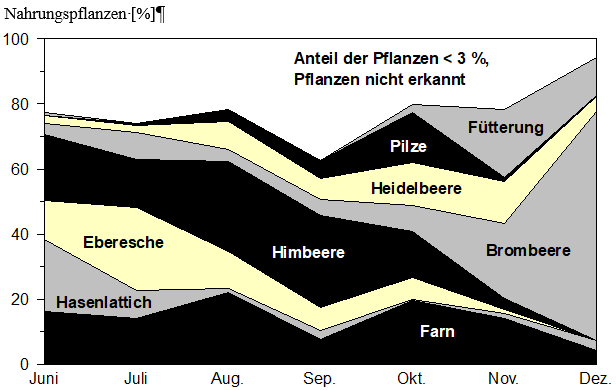
Abb. 4: Durchschnittliche Nahrungszusammensetzung in Rehpansen aus Bodenmais, im Zeitverlauf (n = 114). Untersuchungszeitraum Juni – Dezember, 1987 – 1990
Saisonale Nahrungswahl
Blätter von Hasenlattich und Eberesche wurden besonders häufig im Juni/Juli von den Rehen gefressen, in den nachfolgenden Monaten nahm ihre Bedeutung als Äsungspflanzen ab. Dagegen wurden Himbeerblätter kontinuierlilch den ganzen Sommer über geäst, erst nach dem Absterben der Blätter im Oktober/November ging der Konsum zurück. Pilze wurden in geringem Maße im August und September, im wesentlichen aber zur Haupt-fruktifikationszeit im Oktober gefressen, wobei ihr Anteil am Nahrungsspektrum 15 % betrug. Blätter der Brombeere wurden den Sommer über nur selten gefressen.
Im November/ Dezember waren sie dagegen die wichtigste wildwachsende Nahrung. Dies hängt vermutlich damit zusammen, daß ihre Blätter wintergrün sind und sich, zumindest zu Beginn des Winters, oberhalb der Schneedecke befinden. Farne waren in der Ernährung der Rehe von Juni bis Dezember gleichbleibend von großer Bedeutung. Ihr Anteil am Nahrungs-spektrum war im August, mit 22 % und im Oktober, mit 20 % besonders hoch. Ab Oktober, je nach Schneelage, begannen die Rehe auch Nahrung an den Fütterungen anzunehmen.
Nach Hypogäen wurde bei den Magenanalysen nicht explizit gesucht. Eventuell nehmen Rehe Hirschtrüffel als Aphrodisiakum auf, Arbeiten von Pokorny et al. (2007) deuten darauf hin.
Individuelle Vorlieben
Bei den Untersuchungen stellte sich heraus, daß einige Pansen Pflanzenarten enthielten, die in anderen Pansen nicht nachweisbar waren, dabei handelte sich um Sauerampfer (Rumex spec.), Faulbaum (Frangula alnus), Heidekraut (Calluna vulgaris) Wald-Erdbeere (Fragaria vesca) und Frauenmantel (Alchemilla spec.). Weitere 10 Pflanzenarten kamen nur in zwei oder 3 Pansen vor. Ein Teil der Rehe hatte wohl individuelle Vorlieben für bestimmte Nahrungspflanzen.
Unterschiede in der Zusammensetzung des Artenspektrums in den Pansen könnten allerdings auch durch die unterschiedliche Abundanz einzelner Arten im Untersuchungs-gebiet bedingt sein. Aufgrund der großen Höhenunterschiede im Untersuchungsgebiet
(400 m bis 1.400 m), kommen einige Pflanzenarten in den oberen Höhenzonen nicht mehr vor bzw. umgekehrt. Ein Beispiel dafür ist der Dornfarn, dessen obere Verbreitungsgrenze bei etwa 1.100 m u. M. liegt. Des weiteren beeinflußen natürlich kleinräumige Änderungen in den Standortbedingungen die Verbreitung einzelner Pflanzenarten.
Vergleichbarkeit der Ergebnisse
Die Ergebnisse geben das Nahrungsspektrum von Rehen aus Bodenmais für den Zeitraum von 1987 – 1990 wieder. Es ist anzunehmen, das sich die qualitative Zusammensetzung der Nahrung sowie die Bedeutunug der wichtigsten Äsungspflanzen der Rehe auch in anderen Jahren nicht wesentlich ändert. Dagegen könnten die mengenmäßige Anteile einzelner Pflanzenarten am Nahrungsspektrum in anderen Jahren durchaus variieren. Dies hängt damit zusammen, daß die Zusammensetzung der Waldbodenvegetation nicht statisch sondern dynamisch ist. So ändert sich z.B. nach einem Kahlschlag aufgund des nunmehr höheren Lichtflusses auf den Waldboden die dortige Pflanzengesellschaft beträchtlich.
Auch sogenannte Mastjahre, wie von Bucheckern und Pilzen bekannt, dürften die Nahrungswahl der Rehe beeinflussen. Der relativ hohe Pilzanteil an der Nahrung der Rehe von durchschnittlich 15 % im Oktober während des Untersuchungszeitraums, dürfte z.B. das Ergebnis von mehrern, pilzreichen Jahren gewesen sein. In den nachfolgenden zwei Jahren waren im Untersuchungsgebiet nur wenig Fruchtkörper von Pilzen zu finden und ihr Anteil am Nahrungsspektrum der Rehe vermutlich deutlich geringer.
Deutliche Unterschiede in den Cs-137 Gehalten bei Rehwild zwischen Untersuchungsgebieten
Für den gesamten Untersuchungszeitraum, von 1987-2001, ergibt sich für Cs-137 in Rehen (n= 1.460) eine effektive Halbwertzeit von 6 Jahren. (P<0.0001). Dieser Wert entsteht aus der Kombination von zwei Phasen mit sehr unterschiedlicher Dynamik: In der ersten Phase, von 1987 bis 1995, erfolgt ein relativ schneller Rückgang der Aktivität, mit einer Halbwertzeit von 4,6 Jahren (n=1.091, P<0.0001), während von 1996 bis 2000, mit einer Halbwertzeit von -89 Jahren (n=338, P=0.8454) eine deutliche Verlangsamung eintrat. Obwohl dieser Wert nicht signifikant ist und durch die ausnehmend hohe Kontamination der Rehe im Jahr 2000 besonders beeinflusst wird, besteht seit 1996 der Trend zu größeren, effektiven Halbwertzeiten.
Aus dem Untersuchungsgebiet Göttingen wurden im Oktober 2000, dem Monat in dem die Cs-137 Aktivität von Rehen normalerweise am höchsten ist, 4 Proben untersucht. Alle Proben enthielten weniger als 0,5 Bq/kg. Die Cs-137 Kontamination von Rehen, wie auch von Wildschweinen, variiert innerhalb der Bundesrepublik um 4 Größenordnungen.
Literatur
Die Angaben stammen aus dem Forschungsvorhaben „Untersuchungen zum Verhalten von Radiocäsium in Wildschweinen und anderen Biomedien des Waldes“ BMU 205-675
Die Arbeiten wurden mit Mitteln des Bundesministeriums für Umwelt, Naturschutz und Reaktorsicherheit finanziert. Der Text gibt die Auffassung und Meinung des Auftragnehmers wieder und muss nicht mit der des Bundesumweltministeriums übereinstimmen.
Albers U., 1990: Botanische Panseninhaltsanalyse bei Rehwild im Staatlichen Bayerischen Forstamt Bodenmais. Diplomarbeit, Forstwissenschaftliche Fakultät, Universität Göttingen.
Chamrad A.D., Box T.W., (1964): A point frame for sampling rumen contents. J. Wild. Mgmt. 28 (3): 473-477.
Bubenik A.D., 1984: Ernährung, Verhalten und Umwelt des Schalenwildes. BLV Verlagsgesellschaft.
Fielitz, U., Klemt, E., Strebl, F., Tataruch, F., Zibold, G., 2009. Seasonality of 137Cs in roe deer from Austria and Germany. J. Environm. Radioactiv. 100:241-249.
Drozdz A., Osiecki A., 1973: Intake and digestibility of natural feeds by roe-deer. Acta Theriologica 18: 81-91
Ellenberg H., 1978: Zur Populationsökologie des Rehes (Capreolus capreolus L., Cervidae) in Mitteleuropa. Spixiania 2.
Klötzli F., 1965: Qualität und Quantität der Rehäsung. Verlag Hans Hübner, Bern.
Pokorny, Boštjan; Zaluberšek, Meta; Al Sayegh Petkovšek, Samar.
Scraping of roe deer in forested habitats in early spring: A marking behaviour or searching for hypogeous fungi (Elaphomyces sp.) as an additional food source? // Second International Symposium ‚Game and Ecology‘ / Janicki, Zdravko, editor(s).
Zagreb : Zavod za biologiju, patologiju i uzgoj divljači, Veterinarski fakultet, 2007. 74-75 (poster,domestic peer-review,abstract,scientific). Link